Sexual Reproduction vs. Cloning
The persistence of sexual reproduction among eukaryotes remains a central enigma in evolutionary biology, primarily due to the significant demographic and energetic costs associated with the maintenance of males. In most multicellular organisms, the biological objective of propagating genetic material is achieved through a complex, biparental process that involves the reduction of the genome via meiosis and its subsequent restoration through syngamy.
When contrasted with parthenogenesis—a uniparental form of reproduction where offspring develop from unfertilized eggs—sexual reproduction appears remarkably inefficient. Parthenogenesis allows for a much faster rate of population growth because every individual in a clonal lineage is capable of producing offspring, circumventing the “two-fold cost of males” that halves the reproductive potential of sexual populations.
However, the overwhelming prevalence of sex across the eukaryotic tree of life suggests that the genetic variation it generates acts as a “path to perpetuity,” providing a mechanism for long-term survival that clonal lineages lack. This report evaluates the evolutionary advantages of the “genetic lottery” inherent in sexual reproduction, focusing on its role in environmental adaptation, the purging of deleterious mutations, and the avoidance of extinction over geological timescales.
Comparative Framework of Reproductive Strategies
To understand why sexual reproduction facilitates long-term lineage persistence, it is necessary to contrast its fundamental features with those of parthenogenesis. The following table summarizes the primary differences in genomic handling and evolutionary potential between these two modes of reproduction.
| Feature | Sexual Reproduction | Parthenogenesis (Asexual) |
|---|---|---|
| Source of DNA | Biparental (Two parents) [1] | Uniparental (One parent) [2] |
| Genetic Variation | High (Crossing over + Independent Assortment + Fusion) [3] | Low (Clonal or semi-clonal; relies on rare mutation) [4] |
| Mutation Handling | Can “purge” deleterious mutations through recombination [5] | Mutations accumulate over time (Muller’s Ratchet) [6] |
| Evolutionary Speed | Fast; high adaptability to changing conditions [3] | Slow; relies on sequential accumulation of random mutations [7] |
| Reproductive Potential | Lower (Two-fold cost of males) [8] | Higher (Twice as efficient in “making babies”) [9] |
| Mechanism | Meiosis and Syngamy [1] | Mitosis or modified Meiosis (Apomixis/Automixis) [2] |
While parthenogenesis offers a short-term advantage in population expansion, the lack of genetic mixing creates a rigid genomic structure. This rigidity leaves clonal lineages vulnerable to environmental shifts and the relentless accumulation of genetic errors, eventually leading to their status as “evolutionary dead ends.”
The Mechanistic Foundation of the Genetic Lottery: Meiosis and Recombination
The genetic lottery is not a metaphorical concept but a biological reality facilitated by the intricate molecular mechanisms of meiosis. Unlike mitosis, which preserves the parental genotype in daughter cells, meiosis is designed to maximize diversity through three primary points of randomization: crossing over, independent assortment, and random fertilization. [10]
Molecular Dynamics of Crossing Over
During the first stage of meiosis, Prophase I, homologous chromosomes—one inherited from each parent—pair up to form bivalents or tetrads. [1] This physical alignment allows for the exchange of genetic material between non-sister chromatids, a process known as crossing over or recombination. [3] At specific points called chiasmata, the DNA strands of homologous chromosomes break and rejoin with their counterparts, creating recombinant chromosomes that possess a unique mosaic of alleles that never existed in either parent. [11]
The molecular machinery involved in crossing over is closely linked to the cell’s systems for repairing DNA damage. Many of the tools used to facilitate homologous recombination during meiosis are the same enzymes responsible for repairing double-strand breaks in somatic cells. [12] This suggests that one of the ancestral functions of sex may have been the immediate repair of genomic damage, providing an “undamaged” template for the next generation. [13] In a sexual population, usually one to three crossover events occur per chromosome, ensuring that linkage groups are constantly broken and rearranged. [14]
The Combinatorial Explosion of Independent Assortment
The second mechanism of the genetic lottery occurs during Metaphase I, where the law of independent assortment dictates the random orientation of homologous pairs along the cell’s equator. [1] The orientation of one pair is entirely independent of the others; for example, a maternal chromosome 1 may segregate to the same pole as a paternal chromosome 2. [15] For a diploid organism with \$n\$ pairs of chromosomes, the number of possible gametic combinations is 2 USD^n. USD [16]
In humans, where \$n = 23\$ [12], the potential for diversity is mathematically immense. A single individual can produce 2 USD^{23}\$ or approximately 8,388,608 unique combinations of maternal and paternal chromosomes. [1] When combined with the effects of crossing over, the number of unique gametes produced by a single human is effectively infinite. [14] Random fertilization further amplifies this diversity; the fusion of any genetically unique sperm with any unique egg results in a zygote that represents one out of trillions of possible genetic outcomes. [10]
The Lottery Principle and Adaptive Landscapes
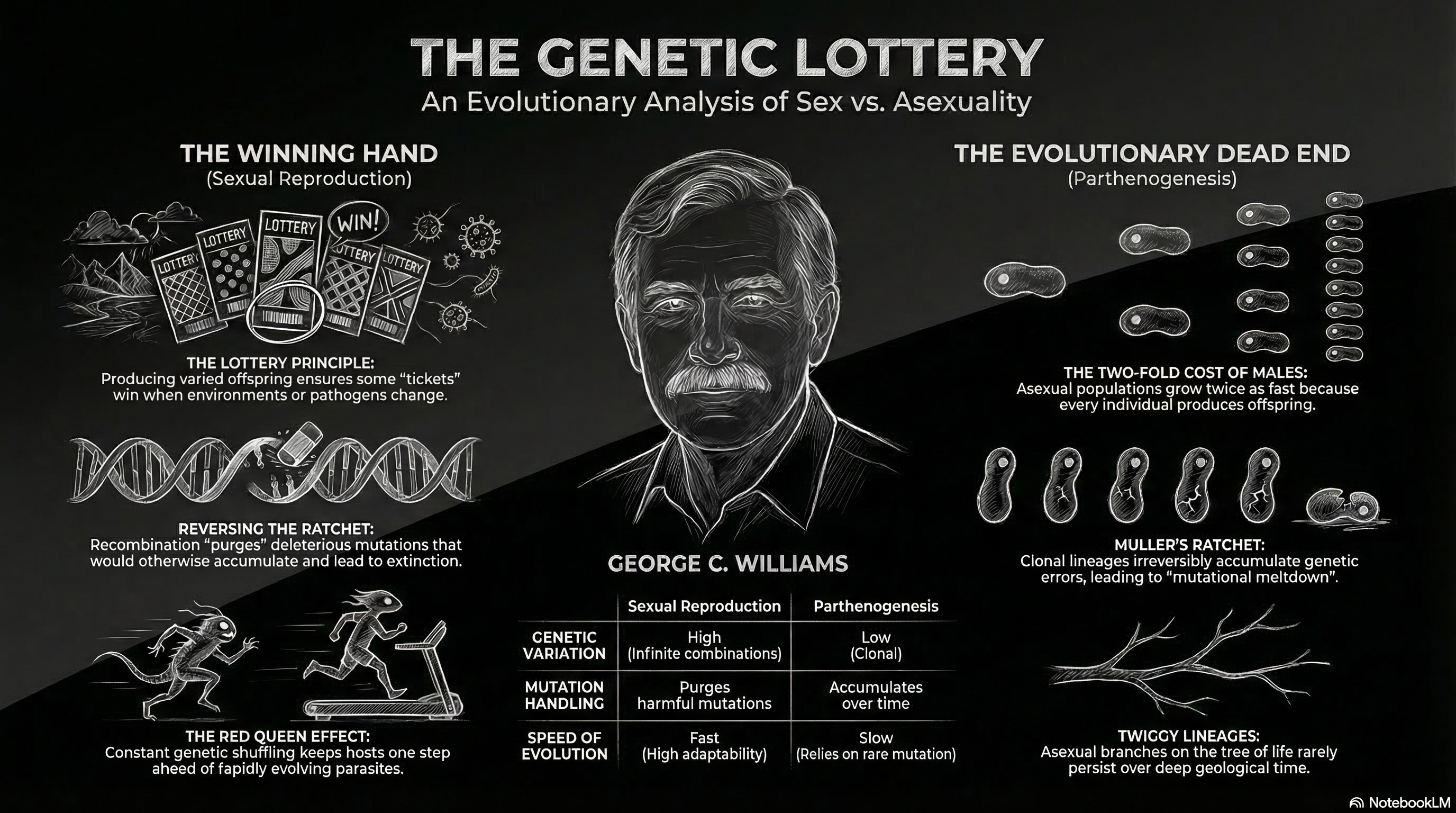
The “Lottery Principle,” first proposed by George C. Williams, provides the theoretical framework for why this variation is favored by natural selection. Williams utilized a lottery analogy to illustrate the disadvantage of asexuality in unpredictable environments: reproducing asexually is equivalent to buying a large number of lottery tickets that all share the same number. [4] If the environment changes such that the parent’s genotype is no longer a “winning combination,” the entire clonal lineage fails. [11]
In contrast, sexual reproduction is akin to purchasing a smaller number of tickets but ensuring that each ticket has a different number. [2] This “bet-hedging” strategy ensures that at least some offspring will possess the phenotypic traits necessary to survive under novel conditions. [7] While this model predicts that sex should be more prevalent in unstable or transient habitats, empirical evidence has shown that sex is actually highly common in stable environments like the tropics, suggesting that biological complexity (such as interaction with pathogens) creates a form of “intrinsic instability” that necessitates constant variation. [4]
Mutation Management and Genomic Purging: Reversing the Ratchet
One of the most significant arguments for sexual reproduction as a “path to perpetuity” involves its capacity to handle deleterious mutations. In the absence of recombination, asexual lineages are subject to an irreversible accumulation of harmful genetic changes known as Muller’s Ratchet. [6]
Muller’s Ratchet and the Genetic Load
In a finite population of asexual organisms, the “least-loaded” class—those individuals with the fewest deleterious mutations—is vulnerable to loss through random genetic drift. [6] Because these organisms inherit their genomes as indivisible blocks, they have no mechanism to “clean” the genome. Once the mutation-free class is lost, it cannot be recovered (except by rare back-mutation), and the minimum mutational load of the population increases. [6] This process acts like a ratchet, clicking forward and irreversibly increasing the “genetic load” of the lineage. [6]
Over time, the accumulation of mutations results in a decline in mean fitness, which can lead to a “mutational meltdown.” [6] As fitness declines, the population size shrinks, making the population even more susceptible to genetic drift and further “clicks” of the ratchet, ultimately resulting in extinction. [6] Laboratory experiments have confirmed this meltdown in RNA viruses and bacteria under conditions where recombination is suppressed. [6]
Synergistic Epistasis and the Deterministic Mutation Hypothesis
The “Deterministic Mutation Hypothesis,” championed by Alexey Kondrashov, suggests that sex is favored if deleterious mutations interact synergistically—meaning each additional mutation causes a disproportionately larger decline in fitness than the one before. [17] Recombination breaks up the linkage between these mutations, concentrating many harmful alleles into specific “severely unfit” individuals. [13] These individuals are then removed from the population by natural selection, effectively purging the population of multiple deleterious mutations simultaneously. [18]
By “shuffling the deck,” sexual reproduction essentially rotates the ratchet backward, allowing offspring to be produced with fewer mutations than their parents. [9] This purging mechanism is essential for the long-term maintenance of complex genomes, which are otherwise destined for degradation in the absence of recombination. [19]
The Coevolutionary Arms Race: The Red Queen Hypothesis
While mutation purging provides a long-term advantage, the Red Queen Hypothesis explains the immediate, short-term maintenance of sex through the lens of host-parasite coevolution. [20] Parasites and pathogens, characterized by short life cycles and rapid evolution, are constantly evolving to “pick the locks” of their hosts’ immune systems. [9]
Parasites as Drivers of Variation
The Red Queen Hypothesis posits that sex is beneficial because it enables hosts to rapidly evolve new genetic defenses against parasites that would otherwise adapt to infect the most common genotypes in a population. [21] In an asexual population, every individual shares the same immune “signature,” allowing a specialized parasite to potentially wipe out the entire lineage. [9] By producing genetically unique offspring, sexual reproduction ensures that the host population remains a “moving target.” [21]
| System | Evidence for Red Queen Hypothesis |
|---|---|
| Potamopyrgus antipodarum (Snail) | Higher frequency of males (sexuals) in shallow waters where parasite-carrying ducks feed. [22] |
| Desert Topminnows (Mexico) | Clonal minnows had significantly more “black spots” from parasites than sexual counterparts in the same pools. [22] |
| C. elegans (Roundworm) | Self-fertilizing populations went extinct when exposed to coevolving bacteria; sexual populations survived. [13] |
| Atlantic Salmon | Wild fish (with mate choice) had higher offspring diversity and fewer roundworm parasites than hatchery fish. [22] |
Empirical tests of the Red Queen Hypothesis have consistently shown that sex is favored in environments where the risk of infection is highest. [22] In the Potamopyrgus snail system, common clonal genotypes were observed to dwindle in number over time as parasites adapted to infect them, while sexual snails persisted by constantly regenerating rare genotypes. [21] This dynamic maintains sexual reproduction even in the face of its two-fold cost, as the fitness benefit of surviving a parasitic infection outweighs the demographic advantage of cloning. [20]
Overcoming Interference: The Hill-Robertson Effect and Adaptive Speed
The “genetic lottery” of sex also resolves a major obstacle to adaptation known as selective interference or the Hill-Robertson effect. [23] In finite populations, natural selection is often hindered because different beneficial mutations arise in different individuals, causing them to compete against each other for fixation. [24]
Clonal Interference and Sequential Fixation
In an asexual population, if two advantageous alleles (\$A\$ and \$B\$) occur in separate lineages, the only way for a single individual to possess both is for the second mutation to independently occur in a lineage that already carries the first. [25] This is statistically rare, meaning beneficial mutations must usually be fixed sequentially. [26] This “clonal interference” significantly slows the rate of evolution. [27]
Sexual reproduction eliminates this interference through recombination. [26] By crossing over, beneficial alleles from different ancestors can be combined into a single lineage, allowing for the simultaneous spread of multiple advantageous traits. [19] This “Vicar of Bray” effect allows sexual populations to adapt to novel environments much more rapidly than asexual ones. [25]
The Impact of Population Size and Recombination Load
The advantages of the Hill-Robertson effect are most pronounced in finite populations where genetic drift is active. [23] In these settings, drift creates “negative linkage disequilibrium,” where high-fitness alleles are found on different chromosomes more often than expected by chance. [28] Recombination breaks down these negative associations, increasing the variance in fitness and allowing selection to act more efficiently. [28]
However, sex also carries a “recombination load”—the potential to break up successful genetic combinations that have already been favored by selection. [19] The persistence of sex suggests that the benefit of creating novel, high-fitness combinations outweighs the risk of disrupting existing ones. [28]
The Economic Paradox: Reconciling the Two-Fold Cost of Males
To define sexual reproduction as a “path to perpetuity,” one must account for its profound short-term disadvantages. The “two-fold cost of sex,” first identified by John Maynard Smith, arises from the production of males in anisogamous species. [27]
Demographic Disadvantage of Sexual Populations
In a sexual population with a 1:1 sex ratio, 50% of the individuals (males) do not directly bear young. [8] If a sexual female produces two offspring, on average only one is a female who can contribute to the next generation’s growth. [8] An asexual female, however, produces two daughters, each of whom can reproduce. [29] This means that a clonal lineage can double in frequency relative to a sexual lineage every generation. [27]
| Generation | Sexual Lineage (Females) | Asexual Lineage (Females) |
|---|---|---|
| 1 | 1 | 1 |
| 2 | 1 | 2 |
| 3 | 1 | 4 |
| 4 | 1 | 8 |
| 5 | 1 | 16 |
Despite this massive 50% fitness penalty, sexual reproduction remains the dominant mode of life for multicellular eukaryotes. [13] The resolution to this paradox is found in the genetic benefits previously discussed: the two-fold cost is “paid” by the increased survival and adaptability conferred by recombination. [29] Furthermore, some models suggest that under certain ecological conditions—such as K-selection (stable environments with high competition)—the survival advantage of sexually derived offspring is worth more than the sheer volume of asexual progeny. [30]
Other Costs and Constraints
In addition to the demographic cost of males, sexual reproduction involves other significant burdens:
- Cost of Finding a Mate: Individuals must spend energy and time locating partners, and they risk failing to find a mate entirely. [27]
- Risk of Disease: Mating is a primary vector for the transmission of parasites and sexually transmitted infections (STIs). [8]
- Predation Risk: Courtship rituals and signals (e.g., peacock feathers or frog calls) can attract predators. [27]
- Meiotic Errors: The complex process of meiosis is prone to errors like nondisjunction, which causes chromosomal disorders such as Down syndrome. [1]
The maintenance of sex despite these multifaceted costs underscores its essential role in long-term evolutionary stability. [31]
Evolutionary Scandals: Lineages That Defy the Rules
While the biological consensus suggests that asexuality is a path to extinction, a few “ancient asexual scandals” have persisted for millions of years without sexual reproduction. [32]
The Bdelloid Rotifers: Masters of Desiccation and Repair
Bdelloid rotifers have existed as an exclusively asexual group for over 40 million years, diversifying into more than 450 species. [33] They have avoided Muller’s Ratchet and the Red Queen through several unique mechanisms:
- Anhydrobiosis and Wind Dispersal: When threatened by parasitic fungi, bdelloids dry up into an inert form called a “tun” and allow themselves to be carried by the wind to new, uninfected habitats. [33] They can survive longer without water than their enemies, effectively “resetting” the arms race. [33]
- Horizontal Gene Transfer (HGT): Bdelloids acquire and use DNA from their environment (including bacteria, fungi, and plants), which provides the genetic variation normally achieved through sex. [34]
- Efficient DNA Repair: They are highly resistant to ionizing radiation because they can repair double-strand breaks in their DNA, potentially using gene conversion to remove deleterious mutations. [35]
Cryptic Sex and “Whiffs” of Recombination
Other supposed ancient asexuals, like Timema stick insects and Darwinulid ostracods, have recently come under scrutiny. [36] Studies have revealed that some “asexual” Timema species occasionally produce functional males or show genomic evidence of rare recombination. [37] This suggests that even a “whiff” of sex—occasional mating events every few thousand generations—may be enough to purge mutations and allow a lineage to persist without the full burden of constant sexual reproduction. [38] Similarly, the discovery of rare males in Darwinulid ostracods suggests they may not be purely asexual after all. [36]
Lineage Selection and the Persistence of Sexual Reproduction
On a macroevolutionary scale, the distribution of asexual lineages suggests they are “twiggy”—they appear frequently as branches on the tree of life but rarely persist long enough to form deep, diverse clades. [31]
Asexuality as an Evolutionary Dead End
Phylogenetic analyses reveal that while transitioning to asexuality is relatively easy (often requiring only a single mutation to lose sexual function), these lineages have much higher extinction rates than their sexual counterparts. [31] Over geological timescales, asexual lineages suffer from a lack of “structural diversification,” making them more vulnerable to mass extinctions and major ecological shifts. [39]
Species Selection for Sexual Maintenance
Lineage selection maintains sex by favoring species that are “trapped” in a sexual state. [31] In many eukaryotes, sex is so deeply linked to survival structures (like seeds in plants or diapause eggs in rotifers) or developmental processes (like epigenetic imprinting in mammals) that its loss would be fatal. [31]
The species that dominate the biosphere today are those that “could not afford to lose sex,” while those that were able to become asexual have largely vanished due to their inability to adapt or manage their genetic load. [31]
Future Outlook: The Role of Facultative Sex
The most successful long-term strategy may not be obligate sex, but facultative sexual reproduction—the ability to switch between sexual and asexual modes. [40] Many organisms, such as Daphnia or the rotifer Brachionus calyciflorus, utilize this strategy to maximize their demographic potential during favorable periods while retaining the ability to “reshuffle the deck” during times of environmental stress. [11]
Experiments with facultative rotifers have shown that the rate of sexual reproduction increases when populations are adapting to novel environments, as the variation generated by sex quickly assembles well-adapted genotypes. [40] Once the population is well-adapted to the new conditions, the rate of sex declines as the “cost of sex” (breaking up fit combinations) begins to outweigh the benefit of creating novel variation. [40] This flexibility suggests that the evolutionary function of the “genetic lottery” is most critical during transitions and crises, providing the necessary plasticity to bridge the gap between stable environmental states. [40]
Conclusion: The Path to Perpetuity
The evidence synthesized from genetic, ecological, and macroevolutionary studies confirms that sexual reproduction is the primary “path to perpetuity” for complex life. While parthenogenesis provides a powerful short-term advantage in fecundity and resource exploitation, it lacks the mechanisms necessary for long-term genomic maintenance and environmental flexibility.
The genetic lottery, powered by the randomization of meiosis and recombination, ensures that sexual populations hold an effectively infinite number of “tickets” in the gamble for survival. By facilitating the purging of deleterious mutations through mechanisms like Muller’s Ratchet and Synergistic Epistasis, and by enabling the continuous evolution of defenses against pathogens via the Red Queen effect, sexual reproduction protects lineages from the inevitable degradation of a static genome.
In the vast timeline of Earth’s history, the diversity produced by sex is the only trait capable of staying ahead of the “ratchet” of decay and the “queen” of coevolution, ensuring that sexual lineages continue to diversify and persist while clonal counterparts fade into evolutionary obscurity.
References
-
Genetics, Meiosis - StatPearls - NCBI Bookshelf, accessed March 9, 2026, https://www.ncbi.nlm.nih.gov/books/NBK482462/
-
Reproduction - bionity.com, accessed March 9, 2026, https://www.bionity.com/en/encyclopedia/Reproduction.html
-
UNIT V: Reproductive Behaviour Evolution of Sex, accessed March 9, 2026, https://www.hhrc.ac.in/ePortal/Zoology/UNIT%20-%20V%20-Reproductive%20Behaviour-converted.pdf
-
Evolution of Sex and the Lottery Principle - AllAboutScience.org, accessed March 9, 2026, https://www.allaboutscience.org/evolution-of-sex-and-the-lottery-principle-faq.htm
-
PMC4224152 (Sexual reproduction allows recombination to… fitness and avoiding Muller’s ratchet), accessed March 9, 2026, https://pmc.ncbi.nlm.nih.gov/articles/PMC4224152/
-
Muller’s ratchet - Wikipedia, accessed March 9, 2026, https://en.wikipedia.org/wiki/Muller%27s_ratchet
-
Why is sexual reproduction more prevalent than asexual reproduction? : r/evolution - Reddit, accessed March 9, 2026, https://www.reddit.com/r/evolution/comments/1fgga7s/why_is_sexual_reproduction_more_prevalent_than/
-
Evolution of Sex - Zoology 500 D, accessed March 9, 2026, https://www.zoology.ubc.ca/~otto/PopGen500/Discussion2/Overheads.html
-
The sexual evolution | Science | guardian.co.uk, accessed March 9, 2026, https://www.theguardian.com/science/story/0,3605,1590096,00.html
-
How does meiosis contribute to genetic variation? - Patsnap Synapse, accessed March 9, 2026, https://synapse.patsnap.com/article/how-does-meiosis-contribute-to-genetic-variation
-
Reproduction - bionity.com (Williams used lottery tickets analogy), accessed March 9, 2026, https://www.bionity.com/en/encyclopedia/Reproduction.html
-
7.3 Does sex lead to fewer mutations? – The Evolution and Biology of Sex, accessed March 9, 2026, https://open.lib.umn.edu/evolutionbiology/chapter/6-3-does-sex-lead-to-fewer-mutations-2/
-
Evolution of sexual reproduction - Wikipedia, accessed March 9, 2026, https://en.wikipedia.org/wiki/Evolution_of_sexual_reproduction
-
Meiosis and Sexual Reproduction | Interactive Textbooks, accessed March 9, 2026, http://vcell.ndsu.nodak.edu/biology_angela_hodgson/Meiosis-p1-v2-page3.html
-
How Meiosis Produces Variation (A-level Biology) - Study Mind, accessed March 9, 2026, https://studymind.co.uk/notes/how-meiosis-produces-variation/
-
Genetic Variation in Meiosis | Overview & Crossing Over - Lesson - Study.com, accessed March 9, 2026, https://study.com/academy/lesson/how-meiosis-fertilization-promote-genetic-variation.html
-
Kondrashov’s hatchet hypothesis (ResearchGate), accessed March 9, 2026, https://www.researchgate.net/figure/Kondrashovs-hatchet-hypothesis-Deleterious-mutations-are-eliminated-more-rapidly-from_fig5_226537324
-
Kondrashov’s hatchet hypothesis. Deleterious mutations are eliminated… | Download Scientific Diagram - ResearchGate, accessed March 9, 2026, https://www.researchgate.net/figure/Kondrashovs-hatchet-hypothesis-Deleterious-mutations-are-eliminated-more-rapidly-from_fig5_226537324
-
Let’s talk about sex: Exploring evolutionary explanations for the persistence of sexual reproduction in eukaryotes - Bikuben, accessed March 9, 2026, https://bikuben.w.uib.no/lets-talk-about-sex-exploring-evolutionary-explanations-for-the-persistence-of-sexual-reproduction-in-eukaryotes-2/
-
Sexual reproduction persists in nature… Red Queen hypothesis, accessed March 9, 2026, https://www.biorxiv.org/content/10.1101/637413v1.full-text
-
Benefits of Sexual Reproduction Lie in Defense Against Parasites - NSF, accessed March 9, 2026, https://www.nsf.gov/news/benefits-sexual-reproduction-lie-defense-against
-
7.5 Testing the Red Queen Hypothesis – Introduction to the Evolution and Biology of Sex, accessed March 9, 2026, https://open.lib.umn.edu/evosex/chapter/7-5-testing-the-red-queen-hypothesis/
-
The Hill–Robertson Effect and the Evolution of Recombination - PMC1526660, accessed March 9, 2026, https://pmc.ncbi.nlm.nih.gov/articles/PMC1526660/
-
Hill–Robertson effect - Wikipedia, accessed March 9, 2026, https://en.wikipedia.org/wiki/Hill%E2%80%93Robertson_effect
-
Vicar of Bray (scientific hypothesis) - Wikipedia, accessed March 9, 2026, https://en.wikipedia.org/wiki/Vicar_of_Bray_(scientific_hypothesis)
-
Adaptation in Sexuals vs. Asexuals: Clonal Interference and the Fisher-Muller Model - PMC1456838, accessed March 9, 2026, https://pmc.ncbi.nlm.nih.gov/articles/PMC1456838/
-
If asexual reproduction is a more efficient way for assuring lineage, why did life evolve to reproduce sexually? - Reddit, accessed March 9, 2026, https://www.reddit.com/r/evolution/comments/1j3xc0w/if_asexual_reproduction_is_a_more_efficient_way/
-
An Evolving Genetic Architecture Interacts with Hill–Robertson Effect, accessed March 9, 2026, https://pmc.ncbi.nlm.nih.gov/articles/PMC4896203/
-
Evolution - A-Z - Cost of sex, accessed March 9, 2026, https://www.blackwellpublishing.com/ridley/a-z/Cost_of_sex.asp
-
Two-fold cost of sex and r/K selection theory - Biology Stack Exchange, accessed March 9, 2026, https://biology.stackexchange.com/questions/17454/two-fold-cost-of-sex-and-r-k-selection-theory
-
Lineage Selection and the Maintenance of Sex - PMC3688966, accessed March 9, 2026, https://pmc.ncbi.nlm.nih.gov/articles/PMC3688966/
-
(PDF) Description of a new genus… Considerations on the morphological evolution of ancient asexuals - ResearchGate, accessed March 9, 2026, https://www.researchgate.net/publication/279711707_Description_of_a_new_genus_and_two_new_species_of_Darwinulidae
-
PAU17 How to Survive Without Sex for 50 Million Years? Dry Up - Unirioja, accessed March 9, 2026, https://publicaciones.unirioja.es/catalogo/online/pau_ingles/unidades/PAU17B.pdf
-
How to Survive Without Sex for 50 Million Years? Dry Up | National Geographic, accessed March 9, 2026, https://www.nationalgeographic.com/animals/article/10201-rotifer-survives-without-sex
-
Bdelloid rotifers could redefine animal sex (Quanta Magazine), accessed March 9, 2026, https://www.quantamagazine.org/bdelloid-rotifers-could-redefine-animal-sex-20141119/
-
Living males of the ‘ancient asexual’ Darwinulidae (Ostracoda: Crustacea) - PMC1560310, accessed March 9, 2026, https://pmc.ncbi.nlm.nih.gov/articles/PMC1560310/
-
Asexual Insect Accused Of Having Sex! - Defector, accessed March 9, 2026, https://defector.com/asexual-insect-accused-of-having-sex
-
Red Queen hypothesis - Wikipedia, accessed March 9, 2026, https://en.wikipedia.org/wiki/Red_Queen_hypothesis
-
In gene-centric view, what is the benefit of sexual reproduction? : r/evolution - Reddit, accessed March 9, 2026, https://www.reddit.com/r/evolution/comments/l6q3ig/in_genecentric_view_what_is_the_benefit_of_sexual/
-
The Evolution of Sex Is Favoured During Adaptation to New Environments - PMC3341334, accessed March 9, 2026, https://pmc.ncbi.nlm.nih.gov/articles/PMC3341334/
Tips and Donations
If you enjoyed this deep dive, consider supporting the project with a tip in Sats. It’s a simple, global way to support independent research.
To send Sats, you’ll need a lightning wallet.